Интенсификация сельского хозяйства снижает сложность микробной сети и количество ключевых таксонов в корнях
В исследовании сравниваются три системы — интенсивная, No till и органическая.
Авторы: Самиран Банерджи, Флориан Уолдер, Люси Бючи, Марсель Мейер, Ален Ю. Хелд, Андреас Гаттингер, Томас Келлер, Рафаэль Чарльз и Марсель Г.А. ван дер Хейден.
Департамент агроэкологии и окружающей среды, Цюрих, Швейцария, 2019 год.
Связанные с корнем микробы играют ключевую роль в продуктивности растений, что делает их важными игроками в агроэкосистемах. До сих пор очень мало исследований оценивало влияние различных систем земледелия на корневую микробиоту, и до сих пор неясно, влияет ли интенсификация сельского хозяйства на структуру и сложность микробных сообществ. Мы исследовали влияние традиционного, нулевого и органического земледелия на грибковые сообщества корней пшеницы с помощью секвенирования PacBio SMRT на образцах, собранных с 60 сельскохозяйственных угодий в Швейцарии. Органическое земледелие содержало гораздо более сложную грибковую сеть со значительно более высокой связностью, чем традиционные и беспахотные системы земледелия. Обилие ключевых таксонов было самым высоким при органическом земледелии, где интенсификация сельского хозяйства была самой низкой. Мы также обнаружили сильную отрицательную связь ( R 2 = 0,366; P <0,0001) между интенсификацией сельского хозяйства и связностью сети корневых грибов. Появление таксонов лучше всего объясняется уровнем фосфора в почве, объемной плотностью, pH и микоризной колонизацией. Известно, что большинство ключевых таксонов образуют арбускулярные микоризные ассоциации с растениями и относятся к порядкам Glomerales , Paraglomerales.и Диверсиспоралес. Подтверждая это, обилие микоризных грибов в корнях и почве также было значительно выше при органическом земледелии. Насколько нам известно, это первое исследование, в котором сообщается о таксонах микоризных для агроэкосистем, и мы демонстрируем, что интенсификация сельского хозяйства снижает сложность сети и обилие таксонов в корневом микробиоме.
Введение
Интенсификация сельского хозяйства является одной из наиболее распространенных проблем двадцать первого века. Чтобы идти в ногу с постоянно растущим населением, общая площадь обрабатываемых земель во всем мире увеличилась более чем на 500% за последние пять десятилетий, при этом на 700% увеличилось использование удобрений и в несколько раз увеличилось использование пестицидов. Интенсификация сельского хозяйства вызвала широкий спектр экологических проблем, включая низкую эффективность использования питательных веществ, увеличение выбросов парниковых газов, эвтрофикацию грунтовых вод, ухудшение качества почвы и эрозию почвы. Альтернативные системы земледелия, такие как консервационное земледелие (например, нулевая обработка) и органическое земледелие, получили широкое распространение для уменьшения такого неблагоприятного воздействия на окружающую среду. Органические пахотные земли составляют 2,5% всех пахотных земель в Европе и более 3,5% в Швейцарии. Внедрение нулевой обработки во всем мире увеличилось примерно на 233% за последнее десятилетие и составляет более 3% от общей площади пахотных земель в Швейцарии. Эти системы земледелия используются для поддержания экологической устойчивости и экосистемных услуг, а в основе экосистемных услуг лежит вклад микробных сообществ.
Микробные сообщества играют незаменимую роль в экосистемах и оказывают широкий спектр услуг. В агроэкосистемах микробы модулируют ряд процессов, включая круговорот питательных веществ, разложение органического вещества, стабилизацию агрегатов почвы, симбиотические и патогенные взаимодействия с растениями, и тем самым играют существенную роль в продуктивности и устойчивости агроэкосистем. Интенсификация сельского хозяйства с высоким использованием ресурсов и низким разнообразием сельскохозяйственных культур может повлиять на микробиоту, связанную с почвой и растениями, с последующим воздействием на экосистемные услуги. Все более широкое внедрение нулевой обработки почвы и органического земледелия также требует изучения их воздействия на микробные сообщества. Предыдущие исследования, сравнивающие влияние обычного, нулевого и органического земледелия, в основном были сосредоточены на микробиоме почвы, и наше понимание влияния этих систем земледелия на связанную с корнями микробиоту невелико.
Связанная с корнями микробиота играет ключевую роль в определении надземной продуктивности. Беспахотное земледелие может повлиять на архитектуру корней и их распределение в почве с последующим влиянием на пополнение корней микробами. Тем не менее, очень немногие исследования оценивали влияние нулевой обработки почвы на микробные сообщества корней, а те, которые изучали микробиоту корней, были сосредоточены только на корневых бактериях или конкретных группах грибов, включая арбускулярные микоризные грибы (АМФ), с использованием традиционных методов. Кроме того, влияние интенсификации сельского хозяйства на общие сообщества корневых грибов до сих пор плохо изучено. Корень растения содержит разнообразный набор эндофитных грибов, которые образуют симбиотические, паразитарные или патогенные ассоциации и через такие ассоциации играют ключевую роль в разнообразии растений, составе сообщества и производительности. Широко распространенный симбиоз AMF и ряд преимуществ, оказываемых этими грибами, в настоящее время хорошо известны. Кроме того, микоризоподобные эндофиты Piriformospra indica, также способствуют росту растений, стрессоустойчивости и вызывают местную и системную устойчивость к патогенам. Триходерма spp. также было показано, что они растут эндофитно и усиливают рост растений и системную устойчивость к фитопатогенам. Таким образом, структура и состав сообществ корневых грибов играют важную роль в агроэкосистемах, однако влияние интенсификации земледелия на сообщества корневых грибов остается малоизученным.
Структура микробиома оказывает существенное влияние на его функционирование. Однако изучение структуры микробиома непросто, главным образом из-за сложных взаимосвязей между множеством его членов. Сети сосуществования микробов могут раскрыть такие отношения и дать представление о структуре сообщества. Сетевой анализ оказался особенно полезным в последние годы для понимания того, как ассоциации микробов меняются в зависимости от параметров окружающей среды. Сетевые баллы также можно использовать для статистической идентификации ключевых таксонов, т. е. таксонов, которые имеют большое влияние в сообществе. Недавнее исследование показало, что, несмотря на то, что они численно незаметны, таксоны-краеугольные камни обеспечивают большую биотическую связь с сообществом и, таким образом, могут быть индикаторами сдвигов в сообществе и смены состава. Также было замечено, что влияние абиотических факторов и генотипов хозяев на микробиом растений облегчается через ключевые таксоны, а сложность корневой микробной сети связана с выживанием растений. Интенсификация сельского хозяйства может изменить структуру корневой микробной сети и обилие ключевых таксонов, что, в свою очередь, может повлиять на продуктивность сельскохозяйственных культур. Однако до сих пор не было исследовано, различаются ли корневые микробные сети в органическом, природоохранном и обычном сельском хозяйстве. Уместный вопрос заключается в том, могут ли микоризные грибы, которые широко известны своей ролью в продуктивности растений, также выступать в качестве краеугольных таксонов в микробном сообществе.
Здесь мы исследовали влияние систем ведения сельского хозяйства на структуру сообщества грибов, используя новейшее секвенирование PacBio SMRT и сетевой анализ образцов корней пшеницы, собранных на 60 сельскохозяйственных угодьях в Швейцарии. Мы стремились ответить на следующие вопросы: (а) Влияет ли интенсивность земледелия на структуру и состав грибковых сообществ корней пшеницы? (b) Отличаются ли сложность сети и обилие ключевых таксонов между традиционным, беспахотным и органическим земледелием? (c) Какие таксоны действуют как краеугольные камни и каковы движущие силы таких таксонов в корневой микробиоте?
Материал и методы
Выбор места и примерка
Образцы почвы были собраны в начале мая 2016 года с полей пшеницы на 60 сельскохозяйственных угодьях в северо-восточных и юго-западных регионах Швейцарии. Пшеничные поля обрабатывались либо традиционной обработкой почвы, либо традиционной нулевой обработкой, либо органической обработкой отвальным плугом в течение как минимум последних 5 лет. Системы земледелия были распределены поровну в обоих регионах, и каждая система была представлена 20 сельскохозяйственными угодьями, в результате чего в общей сложности было 60 ферм.
На полях, обрабатываемых традиционным способом, применялись пестициды и синтетические удобрения, и они обрабатывались в соответствии с рекомендациями Федерального управления сельского хозяйства Швейцарии « Доказательство экологической эффективности». Поля с нулевой обработкой почвы не обрабатывались, за исключением периодического использования полосной обработки и возможного внесения синтетических веществ. Органически управляемые поля не получали пестицидов и синтетических удобрений и обрабатывались в соответствии с рекомендациями BioSuisse, Федерации швейцарских органических фермеров.
В дополнение к присущим системам земледелия различиям в использовании плуга или синтетических удобрений и средств защиты растений, фермеры также посадили 25 различных сортов пшеницы, все из которых входят в список рекомендуемых сортов озимой пшеницы, ежегодно публикуемый Agrarforschung Schweiz или BioSuisse для традиционных или органически управляемых полей соответственно. В то время как полевые участки показали степень изменчивости текстуры почвы, высоты и среднегодовой температуры, ни один из этих параметров существенно не отличался между системами земледелия. Мы рассчитали индекс интенсивности сельского хозяйства в соответствии с предыдущими исследованиями на основе информации, полученной от 59 фермеров. Индекс интенсивности сельского хозяйства был рассчитан с использованием информации о трех антропогенных факторах воздействия: использование удобрений, использование пестицидов и потребление топлива для сельскохозяйственной техники. Эти факторы также были включены в оценку интенсивности сельского хозяйства в предыдущем исследовании.
На каждом сельскохозяйственном участке было отобрано 18 кернов почвы (диаметром 4 см) на глубине 0–20 см с помощью ручного бура (рис. S2 ).). Эти 18 образцов были смешаны и объединены для получения репрезентативной выборки для фермы. Шнек был очищен между площадками. Пять ненарушенных цилиндрических кернов почвы объемом 100 мл и диаметром 5,1 см были отобраны для измерения объемной плотности, и медиана пяти измерений рассматривалась как оценка объемной плотности для каждого поля. Образцы корней отбирали в июне 2016 г. в период цветения пшеницы (стадия роста 69–75 ВВСН). На каждом участке с помощью вилочной лопаты выкапывали десять растений пшеницы, по пять на разрез. Побеги срезали на высоте ~5 см и все корни определенного участка собирали в полиэтиленовый пакет для последующей обработки. Образцы помещали на лед в холодильник для передачи в лабораторию. Образцы почвы обрабатывали в тот же день, что и сбор, удаляя растительные материалы, гомогенизируют и пропускают через сито с размером ячеек 2 мм. Подобразцы были взяты для различных физико-химических и биологических анализов почвы и хранились при температуре 4 ° C или -20 ° C по мере необходимости.
Анализ растений и почвы
Корневой микробиом включает микробные сообщества, связанные с корнями растений, включая микроорганизмы в эндосфере, ризоплане и ризосфере. Это исследование было специально сосредоточено на корневых эндофитных грибковых сообществах. В лаборатории корни тщательно промывали холодной водопроводной водой. Затем тонкие корни (<1 мм) разрезали на мелкие кусочки длиной около 1 см и тщательно перемешивали. Подобразец из 2 г тонких корней хранили в 1,5 пробирках Эппендорфа, лиофилизировали и хранили при -20 ° C для выделения ДНК. Остальные образцы использовали для определения колонизации АМЖ путем оценки обилия арбускул, гиф или пузырьков по модифицированному методу пересечения линий. Было исследовано не менее 100 пересечений на слайде с двумя техническими повторами с применением слепой процедуры на протяжении всего процесса количественного определения, чтобы избежать субъективности, связанной с происхождением образца. Для образцов почвы измеряли общий фосфор (P), доступный для растений фосфор, pH и объемную плотность с использованием стандартных швейцарских протоколов. Доступный для растений P измеряли в соответствии с Olsen et al. Обилие АМФ в почве оценивали путем экстракции фосфолипидов жирных кислот (PLFA) с последующим анализом на газовой хроматографии и масс-спектрометрии. Мы количественно определили содержание АМФ в почве, используя PLFA 16:1ω5, который считается биомаркером АМЖ, поскольку он составляет большую долю от общего количества PLFA в АМЖ, а также имеет сильную корреляцию между содержанием АМФ в почве и концентрациями PLFA 16:1ω5 наблюдались ранее. Анализ нейтральных липидов или NLFA 16:1ω5 также используется в качестве индикатора биомассы AMF; однако NLFA 16:1ω5 в основном присутствует в органах хранения. Таким образом, это считается слабым индикатором активного AMF в почве, и предыдущее исследование также обнаружило низкое количество NLFA 16:1ω5 в почве.
Экстракция ДНК и секвенирование SMRT
Для каждого образца 200 мг корней (сухой вес) использовали для экстракции ДНК с использованием 600 мл буфера для лизиса NucleoSpin PL1 в течение 15 минут при 65 ° C с последующим использованием набора NucleoSpin Plant II (Macherey & Nagel, Дюрен, Германия). Образцы ДНК амплифицировали с парой праймеров ITS1F-ITS4, нацеленной на всю область ITS (~630 п.н.). Прямой и обратный праймеры были синтезированы с 5-нуклеотидной последовательностью заполнения, за которой следовали метки штрих-кода на 5′-конце, чтобы обеспечить мультиплексирование образцов в рамках одного цикла секвенирования. Подготовку библиотеки и секвенирование SMRT проводили в Центре функциональной геномики Цюриха (http://www.fgcz.ch) на приборе PacBio® RS II (PacBio, Сан-Диего, Калифорния, США). Подробная информация об условиях ПЦР и обработке данных последовательности описана в дополнительной информации. Вкратце, портал SMRT использовался для извлечения циклических консенсусных последовательностей (CCS) из необработанных данных (доступных в Европейском архиве нуклеотидов, номер доступа к исследованию: PRJEB27781). CCS, состоящая как минимум из пяти проходов, дает такой же уровень ошибок, как и платформы для секвенирования 454 или MiSeq. Чтения CCS были отфильтрованы по качеству в Mothur (v.1.35.0). Качественные чтения были демультиплексированы на основе последовательностей праймеров штрих-кода с использованием flexbar. Обнаружение химер de novo выполняли на качественных считываниях с использованием UCHIME. Чтобы избежать нежелательных артефактов с несколькими праймерами, мы удалили чтения, в которых был обнаружен полноразмерный праймер для секвенирования. Мы сгруппировали последовательности качества в операционные таксономические единицы (OTU) при сходстве последовательностей ≥98% с серией сценариев UPARSE. Прочтения были дереплицированы, а единичные и химерные последовательности были исключены для разграничения OTU. OTU с низкой численностью (<0,1% глобальной численности и менее 0,5% численности в конкретной выборке) были удалены из набора данных (рис. S3 ).). Мы нормализовали таблицу OTU, разрежив ее до 1000 чтений на выборку. В среднем было обнаружено 357 OTU на сайт и всего 837 OTU на все 60 сайтов. OTU были классифицированы таксономически по базе данных UNITE . Таблицы OTU и таксономии были отфильтрованы, чтобы исключить OTU, классифицированные как негрибковые.
Статистический анализ
Индексы альфа-разнообразия, такие как богатство OTU, равномерность Шелдона и индекс Шеннона-Уивера, были рассчитаны из таблицы OTU разреженных грибов с использованием пакета phyloseq в R v3.4. Влияние систем земледелия и сортов пшеницы на структуру сообщества грибов оценивали путем выполнения PERMANOVA и канонического анализа главных координат с 999 перестановками в PRIMER-E (PRIMER-E, Плимут, Великобритания). Паттерны бета-разнообразия грибов оценивались только по OTU, которые присутствовали как минимум в двух образцах. Однородность многомерных дисперсий проверяли с помощью теста PERMDISP с использованием матрицы подобия Брея-Кертиса в PRIMER. Мы также определили индикаторные таксоны для каждой системы земледелия, используя функцию « мультипаттерн » впакет indicspecies в R. По сути, этот анализ основан на двух видовых признаках: исключительности (присутствует исключительно в среде обитания) и верности (присутствует во всех образцах этой среды обитания). Значение индикатора рассчитывается на основе этих признаков, чтобы оценить степень, в которой вид является индикатором среды обитания.
Модели совместной встречаемости в грибковых сообществах оценивались путем проведения сетевого анализа с использованием показателей максимального информационного коэффициента (MIC) в статистике MINE. MIC — это информативная оценка, которая выявляет положительные, отрицательные и нелинейные ассоциации между OTU. Сетевой анализ был выполнен на том же наборе OTU, что и тестирование бета-разнообразия, т.е. были включены только те OTU, которые присутствовали как минимум в двух выборках, в результате чего было получено 822 OTU. Общая метасеть была построена из 60 образцов, тогда как три сети для конкретных фермерских хозяйств были построены из 20 образцов в каждой. Ассоциации MIC были скорректированы с учетом частоты ложных открытий (FDR), и окончательные сети были построены со статистически значимыми отношениями ( P < 0,05) после коррекции FDR. Затем сети визуализировали в Cytoscape версии 3.4.0. Для расчета параметров топологии сети использовался инструмент NetworkAnalyzer . Узлы (например, OTU грибов в этом исследовании) являются фундаментальными единицами сети, а ребра представляют соединения или связи между узлами. Таким образом, степень представляет собой количество ребер, соединенных с узлом. Коэффициент кластеризации отражает более высокую связанность между узлами в конкретном регионе сети, тогда как кратчайший путь указывает, насколько быстро информация может перемещаться между двумя узлами. Диаметр сети — это наибольшее расстояние между двумя узлами сети. Мы также оценивали сети по сравнению с их рандомизированными версиями, используя модель Барабаси-Альберта доступен в плагине Randomnetworks в Cytoscape v2.6.1. Узлы в случайной сети могут иметь одинаковое количество степеней, что приводит к распределению Пуассона. С другой стороны, неслучайные сети являются безмасштабными, т. е. распределение степеней показывает степенной хвост, где некоторые узлы показывают более высокие степени, чем остальные. Действительно, структурные признаки сетей корневых грибов, такие как распределение степеней, средний кратчайший путь, коэффициент кластеризации, отличались от случайных сетей с равным количеством узлов и ребер. OTU с наивысшей степенью и наивысшей центральностью близости, а также с наименьшими показателями центральности промежуточности считались ключевыми таксонами. Центральность близости основана на средних кратчайших путях и, таким образом, отражает центральную важность узла в распространении информации. С другой стороны, промежуточная центральность раскрывает роль узла как моста между компонентами сети. Для всей сети в качестве ключевых таксонов были выбраны OTU со степенью выше 50, центральностью близости выше 0,44 и центральностью промежуточности ниже 0,12. Для сетей, специфичных для сельского хозяйства, в качестве ключевых таксонов были выбраны OTU со степенью выше 10, центральностью близости выше 0,28 и центральностью промежуточности ниже 0,18. Мы выбрали единый набор пороговых значений для последовательного сравнения сетей, специфичных для фермерских хозяйств. Мы также рассчитали пропорциональное влияние различных порядков грибов в структуре сети, разделив количество узлов, принадлежащих определенному порядку, на количество общих соединений (ребер). Мы оценили разницу между сетями, специфичными для сельского хозяйства, путем начальной загрузки атрибутов узла (степень, между центральностью и центральностью близости) с 10 000 итераций. Затем мы выполнили тест Колмогорова-Смирнова с двумя выборками, чтобы сравнить атрибуты узлов между сельскохозяйственными системами, используя функцию ks.test , встроенную в пакет статистики в Р. Тест Колмогорова-Смирнова сравнивает общую форму кумулятивного распределения двух переменных, где нуль Гипотеза состоит в том, что переменные имеют одинаковые модели распределения. Для каждой сети атрибуты узла вычислялись методом самонастройки с 10 000 итераций. Кроме того, для вычисления атрибутов узла для каждой фермы мы использовали функцию подграфа в igraph.пакет.
Наконец, мы провели анализ случайного леса, чтобы изучить детерминанты выявленных таксонов краеугольных камней. Random Forest — это мощный инструмент машинного обучения, который обеспечивает высокую точность прогнозирования за счет использования ансамбля деревьев решений на основе выборок из набора данных. Это непараметрический и нелинейный статистический метод, который не имеет предварительных предположений о распределении. Часть набора данных, включенная в выборку, называется данными в пакете , тогда как данные, которые не были взяты, называются данными вне пакета. Деревья полностью выращены для прогнозирования данных вне пакета , а важность конкретной переменной-предиктора получается путем случайной перестановки значений этой переменной для данных из пакета.данные и расчет увеличения среднеквадратичной ошибки. Каждый узел дерева решений связан с подмножеством случайных точек данных из исходного набора данных, и, таким образом, увеличение чистоты узла (что в основном представляет собой уменьшение примеси узла или скорости ошибочной классификации) указывает на важность предикторной переменной. Анализ случайного леса был выполнен с 999 перестановками с использованием пакетов randomforest и rfPermute. Лучшие предикторы были идентифицированы на основе их важности с использованием функций Important и varImpPlot . Повышение чистоты узла и значения среднеквадратичной ошибки использовались для определения значимости предикторов с использованием randomForestExplainer.пакет . Факторы, значимые при P < 0,01, были выбраны в качестве предикторов ключевых таксонов.
Полученные результаты
Общая структура и совпадение
Показатели альфа-разнообразия сообществ корневых грибов существенно не различались между традиционной, нулевой и органической системами. Это также верно для общего таксономического состава. Тем не менее, системы земледелия значительно повлияли на структуру сообщества корневых грибов с тремя отдельными кластерами для полей с органической, традиционной и нулевой обработкой. Тест PERMANOVA также подтвердил значительное влияние систем земледелия (псевдо F = 1,42; P <0,05; объясненная вариация = 4,17%). Незначимый тест PERMDISP ( F = 2,072; P = 0,202) указывает на однородное распределение образцов по системам. Кроме того, попарное сравнение в PERMDISP показало, что не было существенной разницы в дисперсиях между органическим и обычным ( F = 1,068; P = 0,372) и органическим и нулевым посевом ( F = 0,870; P = 0,435). Мы не обнаружили влияния сортов пшеницы на структуру сообщества, и это было подтверждено незначимым тестом PERMANOVA (псевдо F = 0,972; P = 0,595). Однако географическое расположение, то есть северо-восточные и юго-западные регионы, оказало влияние на структуру сообщества корневых грибов. Анализ видов-индикаторов был проведен для проверки того, какие таксоны характерны для каждой из трех систем земледелия. Корень, населяющий Trichoderma , представитель Hypocreales , был единственным индикаторным таксоном для традиционной системы земледелия, тогда как семь таксонов грибов (например, Cyphellophora, Myrmecridium, Phaeosphaeria, Cadophora, Pyrenochaeta, Solicoccozyma и Conocybe ) были индикаторными таксонами для нулевой обработки почвы. Шесть таксонов Sordariales, Cantharellales и Agaricales были индикаторными таксонами для органического земледелия, а Chaetomium и Psathyrella были единственными известными родами.
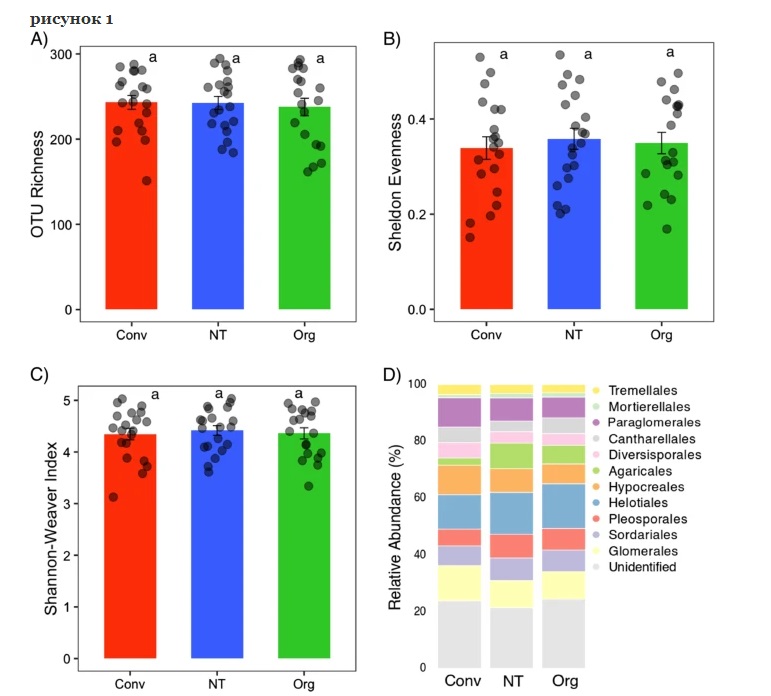
Индексы альфа-разнообразия и состав сообществ корневых грибов в традиционных (Conv), нулевой (NT) и органических (Org) системах земледелия. Богатство OTU ( а ), равномерность по Шелдону ( b ) и индекс Шеннона-Уивера ( c ) были рассчитаны по таблице OTU разреженных грибов. Одна и та же строчная буква указывает на отсутствие статистически значимой ( P <0,05) разницы между системами земледелия. d Гистограмма с накоплением, показывающая относительное обилие сообществ корневых грибов различных порядков пшеницы.

Канонический анализ главных координат (CAP), выявляющий значительное влияние систем земледелия на структуру сообщества грибов. b Общая сеть сообществ корневых грибов в трех системах земледелия. Общая сеть устроена по заказам. Белые, красные и волнистые линии обозначают положительные, отрицательные и нелинейные отношения соответственно. Крупные ромбовидные узлы указывают таксоны краеугольных камней в сети. Десять лучших узлов с наивысшей степенью, наивысшей центральностью близости и наименьшей центральностью промежуточности были выбраны в качестве ключевых таксонов. Из десяти ключевых таксонов в общей сети семь принадлежали к микоризным порядкам, Glomerales, Paraglomerales и Diversisporales .
Сети совместного появления для конкретных фермерских хозяйств
Из-за значительных различий в структуре грибковых сообществ в трех системах земледелия мы дополнительно оценили сети корневых грибов для каждой системы земледелия отдельно. Сети показали значительные различия в их структуре и топологии. Сеть традиционного земледелия состояла из 261 узла (например, таксонов) и 315 ребер (ассоциаций между таксонами), тогда как сеть нулевой обработки состояла из 267 узлов и 341 ребра. В противоположность этому, сеть органического земледелия состояла из 301 узла и 643 ребер. Среднее количество соседей и коэффициент кластеризации сети органического земледелия также были значительно выше, чем для двух других сетей. Более высокая сложность и связанность в сети органического земледелия поддерживались обилием ключевых таксонов. В сети органического земледелия было 27 таких ключевых таксонов по сравнению с двумя в сети с нулевой обработкой почвы и ни одним в традиционной. Большинство этих ключевых таксонов принадлежало к отрядам Glomerales , Tremellales и Diversisporales с заметным присутствием таксонов из отрядов Paraglomerales , Sebacinales и Hypocreales. Чтобы изучить важность ключевых таксонов для более высокой сложности сети в органическом земледелии, мы построили органическую сеть без включения ключевых OTU. Органическая сеть, лишенная каких-либо ключевых таксонов, была намного проще и была похожа на обычную сеть и сеть с нулевой обработкой почвы.

Сети корневых грибов, специфичные для системы земледелия. Каждая сеть была создана с использованием образцов корней, собранных с 20 сельскохозяйственных угодий, принадлежащих к этой системе земледелия. Количество узлов, количество ребер, среднее количество соседей и коэффициент кластеризации приведены ниже конкретных сетей. Крупные ромбовидные узлы обозначают таксоны трапецеидальных искажений, тогда как круглые узлы обозначают другие таксоны в сети. Белые, красные и волнистые линии обозначают положительные, отрицательные и нелинейные отношения соответственно. Несмотря на одинаковое количество узлов, в органической сети было в два раза больше ребер и много тесно связанных узлов, чем в обычных и нулевых сетях, в которых преобладали менее связанные периферийные узлы.

Пропорциональное влияние различных отрядов грибов на сложность корневой микробиоты (левая панель). Влияние было рассчитано путем деления количества узлов, принадлежащих к определенному отряду грибов, на количество общих соединений (ребер). Он иллюстрирует заказы, которые демонстрируют максимальные связи между сельскохозяйственными системами и, таким образом, больше всего влияют на структуру сети. Распределение степеней в трех системах земледелия (правая панель с тремя участками). Степень указывает количество ассоциаций, общих для каждого узла в сети. В обычном земледелии количество градусов было ограничено максимум 12 по сравнению с сетью нулевой обработки, которая имела максимум 22 градуса. С другой стороны, в органическом земледелии было много узлов с температурой более 20 градусов.
Драйверы ключевых таксонов
Интенсивность сельского хозяйства значительно ( P <0,05) различалась в трех методах ведения сельского хозяйства, при этом традиционная система была наиболее интенсивной, а органическая — наименее интенсивной. Эта тенденция была противоположной для подключения к сети, представленной степенью узла в трех методах ведения сельского хозяйства. Начальная загрузка сети показала, что связность сети на органических полях была значительно ( P <0,05) выше, чем на традиционных и нулевых полях. Тест Колмогорова-Смирнова показал, что степень узла, центральность промежуточности и центральность близости значительно ( P <0,01) различались между тремя системами фрейминга. Кроме того, подключение к сети было обратно пропорционально индексу интенсивности сельского хозяйства ( R 2 = 0,366; P <0,0001). Количество ключевых таксонов также было выше (27) в сети органического земледелия, чем в сетях с нулевой обработкой (2) и традиционной (0). Случайный анализ леса показал, что содержание фосфора в почве, объемная плотность, pH и микоризная колонизация лучше всего объясняют ( P <0,01) появление таксонов краеугольных камней. Большинство этих параметров также достоверно ( P <0,05) коррелировали с индексами альфа-разнообразия, что указывает на их важность для общих сообществ корневых грибов. Большинство ключевых таксонов принадлежали к микоризным порядкам, а микоризная колонизация корней пшеницы была значительно ( P <0,01) выше на органических полях, чем на обычных полях и полях с нулевой обработкой. В соответствии с этим, количество микоризных PLFA в почве также было значительно ( P <0,01) выше на органических полях по сравнению с обычными полями. Интенсивность земледелия оказала значительное негативное влияние на микоризную колонизацию корней и численность в почве. В совокупности сложность сети корневых грибов, обилие таксонов краеугольных камней и обилие микоризы показали тенденцию, противоположную тенденции интенсификации сельского хозяйства в системах земледелия.
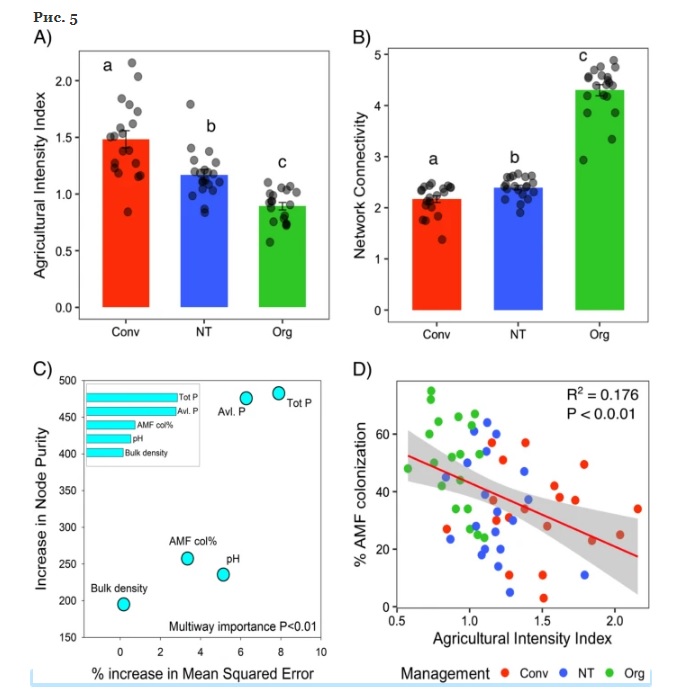
Индекс интенсивности сельского хозяйства для традиционных (Conv), нулевой (NT) и органических (Org) систем земледелия. Индекс интенсивности земледелия рассчитывался с использованием информации о трех антропогенных входных факторах: использовании удобрений, использовании пестицидов и расходе топлива для сельскохозяйственной техники. Различные строчные буквы указывают на статистически значимое ( P <0,05) различие между системами земледелия. b Сетевая связность, представленная степенями узлов для отдельных ферм, рассчитанная путем подмножества сетей трех сельскохозяйственных систем. Различные строчные буквы указывают на статистически значимую (P <0,05) разницу. Результаты случайного анализа леса, показывающие относительный вклад различных факторов в определение численности таксонов краеугольных камней. Среднеквадратическая ошибка (MSE) указывает на точность предсказания каждого фактора. Верхними ( P <0,01) пятью факторами были общий фосфор, доступный для растений фосфор (Olsen P), колонизация корней AMF, pH и объемная плотность. d Взаимосвязь между интенсификацией земледелия и колонизацией корней микоризой. Интенсификация земледелия оказала значимое ( P < 0,01) негативное влияние на корневую колонизацию AMF. Интенсивность сельского хозяйства была самой высокой при традиционном земледелии и самой низкой при органическом земледелии, что было противоположно колонизации AMF.
Обсуждение
В настоящее время хорошо известно, что связанная с корнями микробиота играет важную роль в разнообразии растений, составе сообщества и производительности. Следовательно, важно понять, как сельскохозяйственные методы влияют на микробные сообщества, обитающие внутри корней сельскохозяйственных культур, и как ключевые микробные игроки могут быть нацелены на экологическую интенсификацию агроэкосистем. Однако, поскольку большая часть предыдущей работы была сосредоточена только на микробиоте почвы, наше понимание воздействия систем земледелия на микробиоту, связанную с корнями, все еще находится в зачаточном состоянии. Более того, предыдущие исследования в основном были сосредоточены на моделях микробного альфа- и бета-разнообразия, а влияние различных систем земледелия на структуру микробной сети изучено плохо. Здесь мы показываем, что в корнях пшеницы в разных системах земледелия обитают различные грибковые сообщества и сети различной сложности. Сложность грибковой сети на органически управляемых полях была почти в два раза выше при традиционной и беспахотной агротехнике. Более того, подключение к сети отрицательно ассоциировалось с интенсификацией сельского хозяйства.
Наш вывод о том, что общая структура корневой микробиоты, на которую влияют системы земледелия, согласуется с исследованиями микробиома почвы, в которых большое количество отчетов показало значительное влияние систем земледелия. Следует отметить, что большинство этих исследований изучали микробные сообщества в агрономическом контексте и проводились в полевых условиях. В то время как основная сила полевых испытаний заключается в том, что сельскохозяйственные обработки применяются при однородном управлении и в одном месте с определенным типом почвы, воздействие управления на микробные модели может быть различным на реальных сельскохозяйственных угодьях, и, таким образом, результаты, полученные в одном месте, нельзя обобщать. Мы сообщаем о влиянии методов ведения сельского хозяйства на характеристики корневого микробного сообщества в исследованиях на фермах и во многих областях в региональном масштабе.
Микроорганизмы не развиваются изолированно, а образуют сложные ассоциативные сети. Такие сети имеют особое значение для понимания структуры микробиома и его реакции на факторы окружающей среды. Наше исследование подчеркивает, как методы ведения сельского хозяйства влияют на сетевую структуру корневой микробиоты, и показывает, что органическое земледелие содержит значительно более сложную сеть со многими тесно связанными таксонами (узлами), чем традиционное и беспахотное земледелие. Было показано, что сложные сети с большей связностью более устойчивы к возмущениям окружающей среды, чем простые сети с меньшей связностью. В этом смысле более высокая сложность органических сетей может указывать на то, что корневая микробиота при органическом управлении более устойчива к стрессам окружающей среды, поскольку разные таксоны могут дополнять друг друга. Однако для подтверждения этого наблюдения необходимы дальнейшие исследования.
Keystone таксоны — это таксоны с высокой степенью связи, которые играют важную роль в микробиоме, и их удаление может вызвать значительные изменения в составе и функционировании микробиома. Хотя в предыдущих исследованиях сообщалось о таксонах краеугольных камней в различных средах, сообщения о ключевых таксонах в корневой эндофитной микробиоте очень ограничены. Сеть органического земледелия продемонстрировала самую высокую степень связности и включала большинство основных таксонов. Следует отметить, что грибковое богатство существенно не различалось между системами земледелия, равно как и количество узлов в сетях, специфичных для земледелия, и тем не менее мы наблюдали явную разницу в структуре сети и количестве ключевых OTU. Более того, количество ключевых OTU не различалось между тремя системами земледелия, но эти OTU имели значительно больше ассоциаций с органическим земледелием. Органическая сеть без OTU Keystone была похожа на обычную сеть и сеть с нулевой обработкой почвы, что подчеркивает важность этих элементов для сложности сети. Наши наблюдения показывают, что сложность микробиома не обязательно определяется количеством таксонов в сообществе, а скорее количеством ассоциаций, которые эти таксоны разделяют между собой.
Большинство этих ключевых таксонов были AMF, принадлежащими порядкам Diversisporales, Glomerales и Paraglomerales . Симбиотическую ассоциацию АМП, начавшуюся более 400 млн лет назад, формируют ~80% наземных растений. Наблюдение за тем, что AMF может повышать продуктивность растений, делает их важным игроком в агроэкосистемах. Важность AMF для корневой микробиоты, особенно при органическом земледелии, согласуется с более высоким содержанием AMF в корнях и почвах, наблюдаемым на органических сельскохозяйственных угодьях в этом исследовании. В то время как предыдущие исследования также обнаружили значительно более высокую численность и разнообразие AMF на органических сельскохозяйственных угодьях, чем на обычных, здесь сообщается о важной роли AMF для структуры сети корневых грибов. Один из таксонов немикоризных краеугольных камней в органическом земледелии принадлежал к порядку Sebacinales . Члены этого отряда представляют собой очень разнообразные корневые эндофиты, и считается, что они образуют нейтральные и полезные взаимодействия с растениями. Наше наблюдение за Sebacinales как ключевым таксоном согласуется с предыдущим отчетом, в котором было обнаружено постоянно более высокое обилие Sebacinales на органических сельскохозяйственных угодьях. Поскольку таксоны краеугольных камней связаны со сложностью сети, полезные эндофитные таксоны краеугольных камней, такие как AMF и Sebacinales , могут улучшить сетевое взаимодействие и тем самым усложнить корневой микробиом. Несколько других краеугольных таксонов в общей и органической сетях принадлежали отряду Tremellales . Эта широко распространенная группа базидиомицетов содержит много видов дрожжей и обнаружена в корнях растений в регионах с умеренным климатом. Члены этого отряда грибов также были недавно обнаружены в качестве ключевых таксонов в корневом микробиоме в восьми лесных экосистемах Японского архипелага. Интересно, что мы обнаружили, что два таксона краеугольных камней (OTU_10, OTU_11) были членами Dioszegia.род, который также был обнаружен как краеугольный камень Agler et al. Показано , что влияние абиотических факторов на микробиом опосредуется через Dioszegia у Arabidopsis thaliana . Постоянная идентификация Dioszegia как краеугольного таксона в исследованиях предполагает его важность и подчеркивает потенциал, который можно использовать для манипулирования микробиомом растений. Теперь необходимы дальнейшие исследования, чтобы специально манипулировать этой группой, чтобы проверить, как она влияет на состав и функционирование микробиома. Между таксонами-индикаторами и таксонами-краеугольными камнями не было общих групп грибов. Следует отметить, что таксоны-индикаторы идентифицируются на основе их исключительного обилия (исключительности) во всех пробах (верности) в конкретном местообитании, тогда как таксоны краеугольных камней идентифицируются с использованием сетевого алгоритма, который фокусируется на количестве ассоциаций, которые разделяет OTU, и на его положении в микробиоме. Таким образом, таксоны-индикаторы и таксоны-краеугольные камни отражают два разных микробных индекса, нацеленных на разных членов сообщества.
Важный вопрос заключается в том, как методы ведения сельского хозяйства и интенсивность землепользования влияют на структуру и сложность сети корневых эндофитных грибов? Мы предполагаем, что могут быть два основных механизма: сбор грибов в почве и их рекрутирование и колонизация корней растений. Хорошо известно, что методы ведения сельского хозяйства влияют на качество и количество важных питательных веществ в почве, таких как углерод, азот и фосфор. Сокращенная или нулевая обработка почвы также может изменить объемную плотность верхнего слоя почвы с последующим воздействием на архитектуру корней и удлинение. Эти факторы могут модулировать сборку и эволюцию микробов в почве, тем самым влияя на привлечение микробов в корень. Действительно, мы обнаружили, что уровни фосфора в почве, объемная плотность, а также рН являются определяющими факторами таксонов краеугольных камней, которые связаны со сложностью сети. Большинство таксонов краеугольных камней были микоризными по своей природе, и хорошо известно, что фосфор имеет важное значение для микоризных ассоциаций. Точно так же рН почвы является известной движущей силой грибковых сообществ в почве, особенно микоризных грибов. Таким образом, идентификация характеристик почвы в качестве детерминант ключевых таксонов указывает на важность пополнения как движущей силы сетевой сложности корневой эндофитной микробиоты.
После рекрутирования внутри тела растения микробная адаптация и выживание будут зависеть от физиологических моделей хозяина. Методы ведения сельского хозяйства также могут влиять на физиологические реакции сельскохозяйственных культур за счет наличия воды и питательных веществ, а также применения пестицидов, что может повлиять на поддержание эндофитных микробов в организме растения. Например, известно, что сельскохозяйственные культуры способны снижать выделение углерода микоризным грибам при выращивании в условиях высокой доступности питательных веществ из-за интенсификации сельского хозяйства. Генотипы хозяев также могут влиять на физиологические реакции растений и эндофитную микробиоту, хотя в этом исследовании мы не обнаружили четкой связи между сортами пшеницы и корневыми грибами. Тем не менее, на наших полевых участках росли разные сорта пшеницы, и независимо от того, влияют ли генотипы хозяев на структуру сообщества корневых грибов, потребовался бы эксперимент на конкретном участке с несколькими сортами, выращиваемыми в одних и тех же полевых условиях, что выходило за рамки данного исследования. Предыдущие исследования также показали, что почвенные условия оказывают более сильное влияние на сообщества корневых грибов, чем виды-хозяева, в то время как на сообщества бактерий справедливо обратное. Такие неоднозначные результаты подчеркивают сложную природу взаимодействия растений и микробов и необходимость дальнейших исследований, направленных на изучение факторов, влияющих на эндофитные микробные сообщества сельскохозяйственных культур при различных методах ведения сельского хозяйства. Более того, отбор проб почвы и растений в этом исследовании проводился только в течение одного года, поэтому повторный отбор проб будет следующим шагом для оценки временной согласованности и предсказуемости этих результатов.
Хотя точные факторы сложности сети корневых эндофитов остаются неизвестными, возможно, что состояние питания, обработка почвы, внесение навоза и отсутствие пестицидов могли создать уникальные условия для каждого из трех методов ведения сельского хозяйства, потенциально влияя на сборку грибов в растениях. почве и их поступление в корень растения. Большое количество химических удобрений в традиционной системе земледелия может способствовать развитию быстрорастущих ( r-стратегов ) микробов без сильного давления отбора для какого-либо конкретного таксона и, таким образом, создавая более случайную совокупность. Напротив, применение органических удобрений с более низкой доступностью ресурсов может действовать как селективная сила на сборку грибных сообществ, способствуя медленному росту ( K-стратеги ).) микробы. Вполне возможно, что в микробных сообществах при органическом земледелии могут доминировать К-стратеги , которые медленнее закрепляются и имеют больше шансов на совместное развитие. Для таких микробных сообществ, существующих в условиях ограниченных ресурсов, сотрудничество микробов может быть более важным для выживания. Сотрудничество требует высокой степени связности, что приводит к сетям более высокой сложности. Таким образом, микробные сообщества с более сложной сетью могут быть более распространены при экстенсивном управлении, когда вклады невелики, а ресурсы ограничены, что согласуется с недавним исследованием пастбищ. Количество ключевых таксонов действительно было самым высоким при органическом земледелии, где интенсивность сельского хозяйства была самой низкой, и мы также обнаружили значительную отрицательную связь между интенсификацией сельского хозяйства и сетевой связью. Тем не менее, следует отметить, что микробные таксоны, объединяющиеся в сеть совместной встречаемости, могут быть не связаны с их реальным взаимодействием. Кроме того, в этом исследовании мы рассматривали только корневые грибы, а микробиом включает бактерии, археи и других членов, включение которых необходимо для понимания структуры корневой микробной сети. Также важно отметить, что идентификация таксонов краеугольных камней основана на анализе корреляций (ассоциаций) между таксонами, и необходимы дальнейшие исследования, чтобы показать причинно-следственную связь с точки зрения влияния таксонов краеугольных камней на структуру и функционирование микробиома.
Выводы
Структура и состав корневой микробиоты играют важную роль в агроэкосистемах, однако существует значительный недостаток знаний о влиянии интенсификации сельского хозяйства на корневую микробиоту. Ван дер Хейден и Хартманн подчеркнули важность сетевой структуры для функционирования растительных микробиомов, в то время как Banerjee et al. недавно обобщил ключевые таксоны из различных сред, чтобы подчеркнуть их важность для структуры и функционирования микробиома. Настоящее исследование основывается на этой концептуальной основе и расширяет ее, чтобы продемонстрировать, что интенсификация сельского хозяйства оказывает негативное влияние на структуру сети корневых грибов и обилие таксонов краеугольных камней. Наше исследование показывает, что подключение к сети и обилие ключевых таксонов были самыми высокими при органическом земледелии, где интенсивность ведения сельского хозяйства была самой низкой. Более высокая встречаемость членов микробных сообществ при органическом земледелии может свидетельствовать о большем экологическом равновесии и сложности микробиома, который может быть более устойчивым к стрессам окружающей среды. Ключевым преимуществом этого исследования является то, что образцы были собраны с 60 сельскохозяйственных угодий, и сообщаемые эффекты могут быть обобщены, поскольку образцы были взяты с большого количества полей в разных местах с разными режимами управления. Недавняя концепция умного земледелия (Wolfert et al.) подчеркивает нестандартное мышление . Недавно также был отмечен потенциал использования растительного микробиома для устойчивого сельского хозяйства. Микоризные грибы хорошо известны за их влияние на продуктивность растений, и поэтому таксоны микоризных краеугольных камней могут быть использованы в качестве инструмента для «умного» земледелия.
Источник: https://www.nature.com/articles/s41396-019-0383-2
Союз органического земледелия — независимое общественное движение. Рост производства и потребления здоровых, органических продуктов, обучение, просвещение потребителей, научные исследования, внедрение экоагротехнологий в АПК.
Мы за здоровье почв, экосистем и людей! Вы с нами?
Вступить в Союз органического земледелия
Подписывайтесь на нас в социальных сетях:
Телеграм:
https://t.me/organicsozbio
Вконтакте:
https://vk.com/union_of_organic_agriculture


